为重症救治赋能
为患者康复加速

登录
注册
登录方式
方式一:
PC端网页:www.rccrc.cn
输入账号密码登录,可将此网址收藏并保存密码方便下次登录
方式二:
手机端网页:www.rccrc.cn
输入账号密码登录,可将此网址添加至手机桌面并保存密码方便下次登录
方式三:
【重症肺言】微信公众号
输入账号密码登录
注:账号具有唯一性,即同一个账号不能在两个地方同时登录。
钟佳成,王凌伟 深圳市人民医院呼吸与危重症医学科 深圳市呼吸疾病研究所 深圳市呼吸系统疾病临床研究中心 发布于2023-06-28 浏览 1128
 收藏
收藏
作者:钟佳成,王凌伟
单位:深圳市人民医院呼吸与危重症医学科 深圳市呼吸疾病研究所 深圳市呼吸系统疾病临床研究中心
【摘要】 原发性免疫缺陷病(PID)是一组由遗传缺陷导致免疫系统发育和功能障碍的先天性疾病。患者可出现反复感染、淋巴细胞增殖性疾病、自身免疫性疾病、自身炎症性疾病或恶性肿瘤。呼吸是最常见受累系统之一,包括感染性和非感染性病变。由于这类疾病较为罕见,加之临床医生对其认识尚不充分,很多患者直至成年仍未被诊断。深入了解PID在呼吸系统的表现将有助于提高此类疾病的诊断率。
原发性免疫缺陷病(primary immunodeficiency diseases,PID)又称人类免疫出生错误(human inborn errors of immunity,IEI),通常被认为是由生殖细胞单基因缺陷引起的,表现为严重和/或反复发作的感染性疾病、自身免疫疾病、特应性皮炎以及造血系统和/或实体恶性肿瘤的易感性增加[1]。呼吸系统是PID患者常见的受累部位[2]。本文将综述PID在成人呼吸系统非感染性疾病的表现,旨在提高呼吸科医生对此类疾病的诊断意识。

一、PID的分类
PID可分为以下10种类型[3]:①联合免疫缺陷病:是以T细胞发育或功能障碍为特征的免疫缺陷[4]。②伴有典型症状的免疫缺陷综合征:基因缺陷不仅累及免疫系统,还涉及其他系统。因此在前者的基础上伴随着多种非免疫性的缺陷,如血小板减少症、维生素B12和叶酸代谢缺陷等[5]。③抗体免疫缺陷病(predominantly antibody deficiencies,PADs):由于B细胞缺陷或B细胞与T细胞交互障碍导致的抗体产生受损。由于反复感染,患者常伴随严重的过敏反应和自身免疫性疾病[6],PADs中又以常见变异型免疫缺陷病(common variable immunodeficiency,CVID)为主。CVID是由于B细胞先天缺陷导致的一种或多种免疫球蛋白减少和/或抗体反应受损[7]。④免疫失调性疾病:可表现为自身免疫性疾病(免疫系统过度激活所致的自身反应性淋巴细胞和高滴度自身抗体的产生)、过敏或淋巴细胞增殖[8]。⑤吞噬细胞缺陷病:吞噬细胞功能或数量异常,可导致严重的细菌/真菌感染[9]。⑥天然免疫缺陷病:包含IFN-γ、Toll-IL1R、IRAK1等多种参与天然免疫细胞组分的缺陷[10]。⑦自身炎症性疾病:主要由先天免疫系统紊乱所引起,较少涉及获得性免疫系统中的T细胞或B细胞,其特征是在没有微生物感染的情况下反复发作的发热和全身性炎症[11]。⑧补体缺陷病:维持免疫稳态和免疫监控的补体系统受损,导致全身/局部炎症性和血栓性疾病的风险增加[12]。⑨骨髓衰竭性疾病:包括一系列遗传性和获得性血液病,造成血细胞产生不足,导致严重的并发症,包括贫血、免疫系统功能减弱、凝血功能受损和癌症风险增加[13]。⑩IEI的拟表型是由于体细胞突变或者自身中和抗体造成与胚系IEI类似的表型[14]。
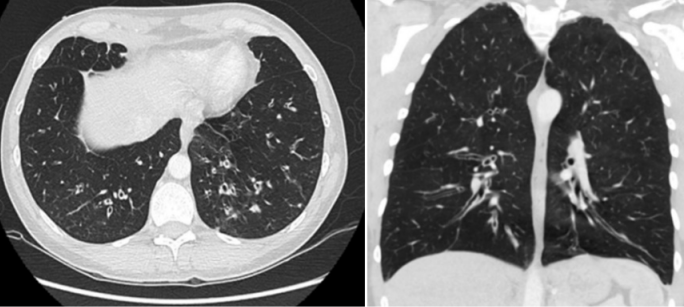
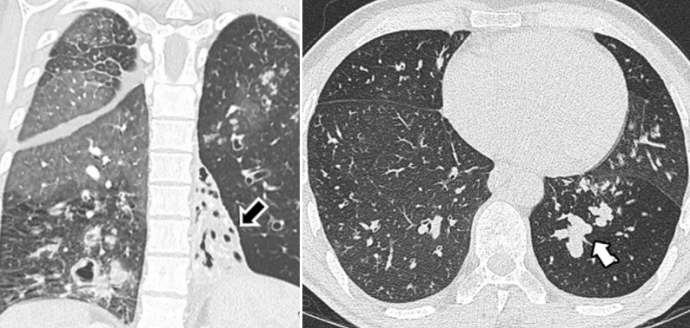
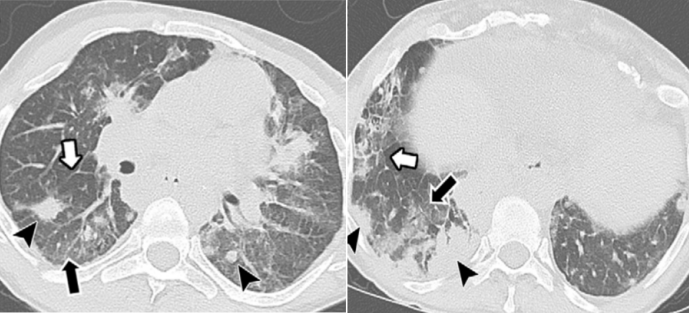
图1 CVID合并哮喘的患者肺部CT:弥漫性气道增厚和支气管扩张,小气道黏液嵌塞,伴远端小叶中心性结节[20]
图片来源:Imaging Features of Primary Immunodeficiency Disorders (DOI: 10.1148/ ryct. 2021200418)
图片来源:Imaging Features of Primary Immunodeficiency Disorders (DOI: 10.1148/ ryct. 2021200418)
三、结语与展望
参考文献
[1] Notarangelo L D, Bacchetta R, Casanova J L, et al. Human inborn errors of immunity: An expanding universe[J]. Sci Immunol, 2020, 5(49):eabb1662.
[2] Hurst J R, Warnatz K. Interstitial lung disease in primary immunodeficiency: towards a brighter future[J]. Eur Respir J, 2020, 55(4):2000089.
[3] 陈然, 罗颖, 杨军, 等. 免疫出生错误分类更新(2022版)解读[J]. 中华儿科杂志, 2022, 60:1262-1265.
[4] Aranda C S, Guimarães R R, de Gouveia-Pereira Pimentel M. Combined immunodeficiencies[J]. J Pediatr (Rio J), 2021, 97 Suppl 1(Suppl 1):S39-S48.
[5] Sharifinejad N, Azizi G, Chavoshzadeh Z, et al. Autoimmunity in monogenic combined immune deficiencies with associated or syndromic features[J]. Front Immunol, 2022, 13:1023127.
[6] Vilela M. Human Inborn Errors of Immunity (HIEI): predominantly antibody deficiencies (PADs): if you suspect it, you can detect it[J]. J Pediatr (Rio J), 2021, 97 Suppl 1(Suppl 1):S67-S74.
[7] Emmaneel A, Bogaert D J, Van Gassen S, et al. A Computational Pipeline for the Diagnosis of CVID Patients[J]. Front Immunol, 2019, 10:2009.
[8] Long A, Kleiner A, Looney R J. Immune dysregulation[J]. J Allergy Clin Immunol, 2023, 151(1):70-80.
[9] Bogaert D J, Haerynck F. Congenital Defects of Phagocytes[M]. In: D'Elios MM, Baldari CT, Annunziato F, eds. Cellular Primary Immunodeficiencies. Cham: Springer International Publishing; 2021:97-176.
[10] Alsina L, Rodriguez-Gallego C, Esteve-Solé A, et al. Defects in Intrinsic and Innate Immunity[M]. In: D'Elios MM, Baldari CT, Annunziato F, eds. Cellular Primary Immunodeficiencies. Cham: Springer International Publishing; 2021:177-212.
[11] Ozkurede V U, Franchi L. Immunology in clinic review series; focus on autoinflammatory diseases: role of inflammasomes in autoinflammatory syndromes[J]. Clin Exp Immunol, 2012, 167(3):382-390.
[12] Schröder-Braunstein J, Kirschfink M. Complement deficiencies and dysregulation: Pathophysiological consequences, modern analysis, and clinical management[J]. Mol Immunol, 2019, 114:299-311.
[13] Malouf C, Loughran S J, Wilkinson A C, et al. Translational research for bone marrow failure patients[J]. Exp Hematol, 2022, 105:18-21.
[14] Tangye S G, Al-Herz W, Bousfiha A, et al. Human Inborn Errors of Immunity: 2022 Update on the Classification from the International Union of Immunological Societies Expert Committee[J]. J Clin Immunol, 2022, 42(7):1473-1507.
[15] Lyons J J, Milner J D. Primary atopic disorders[J]. J Exp Med, 2018, 215(4):1009-1022.
[16] Vaseghi-Shanjani M, Smith K L, Sara R J, et al. Inborn errors of immunity manifesting as atopic disorders[J]. J Allergy Clin Immunol, 2021, 148(5):1130-1139.
[17] Wang Y, Ma C S, Ling Y, et al. Dual T cell- and B cell-intrinsic deficiency in humans with biallelic RLTPR mutations[J]. J Exp Med, 2016, 213(11):2413-2435.
[18] Peled A, Sarig O, Sun G, et al. Loss-of-function mutations in caspase recruitment domain-containing protein 14 (CARD14) are associated with a severe variant of atopic dermatitis[J]. J Allergy Clin Immunol, 2019, 143(1):173-181.e10.
[19] Bjelac J A, Blanch M B, Fernandez J. Allergic disease in patients with common variable immunodeficiency at a tertiary care referral center[J]. Ann Allergy Asthma Immunol, 2018, 120(1):90-92.
[20] Ibrahim H, Walsh J, Casey D, et al. Recurrent asthma exacerbations: co-existing asthma and common variable immunodeficiency[J]. J Asthma, 2022, 59(6):1177-1180.
[21] Brent J, Guzman D, Bangs C, et al. Clinical and laboratory correlates of lung disease and cancer in adults with idiopathic hypogammaglobulinaemia[J]. Clin Exp Immunol, 2016, 184(1):73-82.
[22] Spagnolo P, Fabbri LM, Bush A. Long-term macrolide treatment for chronic respiratory disease[J]. Eur Respir J. 2013 Jul;42(1):239-51.
[23] Khan T Z, Wagener J S, Bost T, et al. Early pulmonary inflammation in infants with cystic fibrosis[J]. Am J Respir Crit Care Med, 1995, 151(4):1075-1082.
[24] Mooney D, Edgar D, Einarsson G, et al. Chronic lung disease in common variable immune deficiency (CVID): A pathophysiological role for microbial and non-B cell immune factors[J]. Crit Rev Microbiol, 2017, 43(4):508-519.
[25] Taneja A, Muco E, Chhabra A. Bruton Agammaglobulinemia. StatPearls. Treasure Island (FL): StatPearls Publishing Copyright © 2023, StatPearls Publishing LLC.; 2023.
[26] Rodriguez J A, Bang T J, Restrepo C S, et al. Imaging Features of Primary Immunodeficiency Disorders[J]. Radiol Cardiothorac Imaging, 2021, 3(2):e200418.
[27] Lopes J P, Ho H E, Cunningham-Rundles C. Interstitial Lung Disease in Common Variable Immunodeficiency[J]. Front Immunol, 2021, 12:605945.
[28] Bates C A, Ellison M C, Lynch D A, et al. Granulomatous-lymphocytic lung disease shortens survival in common variable immunodeficiency[J]. J Allergy Clin Immunol, 2004, 114(2):415-421.
[29] Turner M W. The role of mannose-binding lectin in health and disease[J]. Mol Immunol, 2003, 40(7):423-429.
[30] Garred P, Honoré C, Ma Y J, et al. MBL2, FCN1, FCN2 and FCN3-The genes behind the initiation of the lectin pathway of complement[J]. Mol Immunol, 2009, 46(14):2737-2744.
[31] Beinrohr L, Dobó J, Závodszky P, et al. C1, MBL-MASPs and C1-inhibitor: novel approaches for targeting complement-mediated inflammation[J]. Trends Mol Med, 2008, 14(12):511-521.
[32] Chung W Y, Pollard C A, Stover C, et al. Pilot study: deficiency of mannose-binding lectin-dependent lectin pathway, a novel modulator in outcome from pancreatic islet auto-transplantation[J]. Ann Transl Med, 2020, 8(5):170.
[33] Dicker A J, Crichton M L, Cassidy A J, et al. Genetic mannose binding lectin deficiency is associated with airway microbiota diversity and reduced exacerbation frequency in COPD[J]. Thorax, 2018, 73:510-518.
[34] Tran H B, Ahern J, Hodge G, et al. Oxidative stress decreases functional airway mannose binding lectin in COPD[J]. PLoS One, 2014, 9(6):e98571.
[35] García-García A, Pérez de Diego R, Flores C, et al. Humans with inherited MyD88 and IRAK-4 deficiencies are predisposed to hypoxemic COVID-19 pneumonia[J]. J Exp Med, 2023, 220(5):e20220170.
[36] Zhang Q, Bastard P, Cobat A, et al. Human genetic and immunological determinants of critical COVID-19 pneumonia[J]. Nature, 2022, 603(7902):587-598.
[37] Ivashkiv L B, Donlin L T. Regulation of type I interferon responses[J]. Nat Rev Immunol, 2014, 14(1):36-49.
[38] Bastard P, Rosen LB, Zhang Q, et al. Autoantibodies against type I IFNs in patients with life-threatening COVID-19[J]. Science, 2020, 370(6515):eabd4585.
作者简介
深圳市呼吸疾病研究所副所长
南方科技大学医学院诊断学教研室主任
博士,教授,主任医师,博士生导师
2005年入选中国卫生部国际资源后备人才库人才
2010年德国柏林Evangelische Lungenklinik医院进修
中华医学会呼吸病学分会第十一届委员会委员
中国医学装备协会呼吸病学分会副主任委员
广东省呼吸内科质控中心副主任
深圳市医师协会呼吸医师分会会长
深圳市医学会呼吸病学分会副主任委员
深圳市医防融合项目呼吸项目组组长
科技部重大专项《慢阻肺动物模型病理机制解析及新靶标的发现》课题负责人
声明:
本文仅用于学术内容的探讨和交流,不用于任何商业和推广,亦不作为最终的临床决策。临床实践需根据患者的具体情况选择适宜的处理措施。
 后可发表评论
后可发表评论




友情链接
联系我们
 公众号
公众号
 客服微信
客服微信